meningomyeloseele.fi > tietoa > genetiikka
TIETOA
NEURULAATIO
Koko keskushermosto on peräisin nesteen täyttämän neuraaliputkeksi kutsutun ontelon seinistä. Neuraaliputki muodostuu alunperin ektodermistä peräisin olevasta neuraalilevystä, kun sen keskelle kuroutuu uurre, jonka reunat kallistuvat yhteen ja kiinnittyvät toisiinsa. Hermostoputken sulkeutumisessa on kaksi pääpiirrettä. Itse putken muodostuminen sekä alkuaan hyvin laakean hermostolevyn piteneminen yhdessä koko alkion kanssa. Yleisesti tätä yhteistapahtumaa kutsutaan neurulaatioksi. Ensimmäistä osaa, uurteen muodostumista ja sen yläosien kiinnittymistä, ohjaa monien muiden ohessa indusoivana tekijänä helposti kudoksissa diffundoituva peptidi-kasvutekijä sonic hedgehog (SHH), joka erittyy selkäjänteestä. Hermostolevyn pidentymistä puolestaan ohjaa planaarisen solu-polaarisuuden reitti. Monet kehityksessä havaituista mutanttimuodoista ovat näyttäneet myös olevan paikkaspesifisiä, mikä on johtanut siihen että ennen vallalla ollut Zipper, eli vetoketju-malli, jossa hermostoputken sulkeutuminen tapahtuisi kahdessa vaiheessa, primäärisessä ja sekundäärisessä, on korvattu modernilla mallilla, jossa primaarisen neurulaation alueessa on neljä sulkeutumisen solmukohtaa. Eli neuraaliputki sulkeutuu viidessä vaiheessa. Suurin osa yksilön henkiin jättävistä sulkeumishäiriöistä aiheutuu alueen 1. hännänpuoleisessa päässä.
Takaisin sivun alkuun
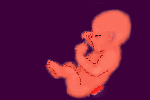
NEURAALIPUTKEN SULKEUTUMISHÄIRIÖT (NTD:T)
Kehitysgeneettisissä häiriöissä on otettava huomioon myös viallisen kudoksen induktiivinen jälkivaikutus. Se että neuruloituminen ei etene normaalisti jossakin kohtaa, vaan jää vajaaksi, katkaisee normaalin induktion kulun myös tästä eteenpäin ja näin myös selkärangan ja halkiolle läheisen lihaksiston ja ihon kehittyminen jää vajaaksi. Hermostoputken sulkeutumishäiriöt (NTD, neural tube defects) tapahtuvat tavallisesti jo hyvin varhaisessa vaiheessa, kun alkion hermoston muodostavan hermostolevyn vaiheittainen sulkeutuminen neuraaliputkeksi häiriintyy syystä tai toisesta.Tämä muutoksille altis yksilönkehityksen vaihe on 22.-28. raskauspäivän aikana, eli varsin varhaisessa vaiheessa, jolloin suuri osa äideistä ei ole edes vielä perillä raskaudestaan. Neuraaliputken yleisesti patologisia sulkeutumishäiriöitä on periaatteessa kolmea tyyppiä. Meningomyeloseele eli MMC, Anenkefalia (aivottomuus) ja enkefaloseele (aivotyrä). MMC on vakavin muoto hermostoputken kaudaalisista halkioista (spina bifida) . Hermostoputken alkupään sulkeutumisen häiriöt anenkefalia ja enkefaloseele puolestaan johtaa vaikeisiin kallon ja aivojen kehityshäiriöihin. Anenkefaliassa kallon yläosa jää kehittymättä ja aivokudoksesta suurin osa tuhoutuu. Tällaiset lapset eivät elävänä syntyessäänkään elä tavallisesti kuin muutaman päivän. Harvinaisemman ja lievemmän enkefaloseelen eli aivotyrän ennuste taas riippuu tyrän sisällä sijainneesta aivomateriaalin määrästä. MMC:ssä osa selkäytimestä ja sitä suojaavista kalvoista jää avoinaisiksi, koska hermostoputken reunat eivät ole kuroutuneet vauriokohdassa kiinni toisiinsa. Tämän seurauksena sillä kohdalla selkärangan takanikamakaaret ovat jääneet sulkeutumatta tai muodostumatta, eikä niiden kohdalle ole yleensä kasvanut normaalisti lihaksia ja ihoa. Sikiön kehityksen jatkuessa selkäydintä ympäröivät kalvot ja itse selkäydin usein pullistuvat selkäydinkanavasta aukkoon muodostaen aivoselkäydinnesteen täyttämän kohjun, joka voi olla joko selkäydinkalvon tai ihon peittämä. Selkäydinkohjuista on olemassa myös lievempiä versioita, joissa esimerkiksi Spina bifida occultassa pieni osa sulkeutumasta on vaillinainen johtaen yleensä varsin vähäisiin vaurioihin läheisissä selkärangan nikamissa. Nämä häiriöt ovat varsin yleisiä ja yleensä niin lieviä että ne jäävät huomaamatta koko kantajan eliniän ajan. Toisinaan niistäkin voi kuitenkin koitua ongelmia selkäytimen liekautumisen johdosta, tavallisesti murrosiässä jos selkäydin tai siitä lähtevät hermot takertuvat kiinni rakennepoikkeaman kohtaan eikä selkäydinpääse liukumaan normaalilla tavalla ylöspäin selkärangan pidentyessä ympärillä.
Takaisin sivun alkuun
NTD:IHIN JOHTAVAT HÄIRIÖT
Pitkään aikaa oltiin sitä mieltä, että hermostoputken sulkeutumishäiriöillä ei olisi geneettistä pohjaa, sillä se ei näytä juuri osoittavan mendelistisen periytymisen pääperiaatteita, lukuunottamatta joitakin harvinaisia esimerkkejä, joihin palaamme myöhemmin. Myös taudin uusiutuvuus on varsin pieni ollakseen geneettinen ominaisuus: Riski että perheeseen, jossa on yksi sairas lapsi syntyisi toinen sellainen on vain 3-5% luokkaa, kun lajimme keskimäärinen frekvenssi maailmassa on 1/1000-luokkaa ja uusiutumisriskiä on havaittu voitavan pienentää suurilla ravinnelisäannoksilla (foolihappoa). Geneettisen periytyvyyden sijaan osoitettiin siis paljon huomiota ympäristön vaikutukseen. Havaittuja riskitekijöitä geneettisen alttiuden lisäksi ovat: useat teratogeeniset aineet ja ympäristössä vaikuttavat tekijät, kuten genotoksiset aineet ja säteily, aineet, jotka näyttävät aiheuttavan spesifisesti selkärankahalkioita, kuten epilepsia-lääke Valproaatti, raskauden kriittisenä aikana koettu kuume tai esim. pitkät kylvyt kuumassa vedessä, ja yllättäen myös monilla varsin tavallisilla ja jopa normaalisti hyödyllisilläkin aineilla on havaittu olevan sulkeutumishäiriöitä tuottavia sivuvaikutuksia, kuten kuten salisylaatin ja A-vitamiinin runsailla annoksilla. Kumminkin on myös selvää tilastotietoa siitä että häiriöiden frekvenssit eri ihmispopulaatioissa vaihtelevat erittäin runsaasti. Esimerkiksi Englannissa erilaiset havaitut neuraaliputken sulkeutumishäiriöt ovat siitä yleisimpiä ja frekvenssi on pelkästään MMC:llä (kun otetaan huomioon myös abortoidut yksilöt) 1/500, kun Suomessa frekvenssi on kaikkien havaittavien NTD:ien suhteen yhteensä luokkaa 7.3/10 000 (Stakes, mukaanlukien abortoidut), ja MMC:llä 1/2500. Neuraaliputken sulkeutumishäiriöiden geneettiset tekijät voivat olla joko polygeenisiä tapahtuman riskiä tilastollisesti kasvattavia häiriöitä, tai jopa yhden mutatoituneen geenin aiheuttamia mendelistisesti periytyviä resessiivisiä virheitä, joista jälkimmäisiä on diagnosoitu ihmispopulaatioista ilmeisesti vain joitakin yksittäisiä kertoja. Harvinaisuus voi hyvinkin johtua siitä, että vaikka aika monen yksittäisen geenin häiriöt voivat hiirimallien mukaan aiheuttaa neuraaliputken sulkeutumishäiriöitä, on tällaisia yksittäisestä geenistä johtuvia ominaisuuksia vastaan populaatioissa voimakasta karsintapainetta vaikka ne olisivatkin resessiivisiä.
Takaisin sivun alkuun
FOLAATIN AINEENVAIHDUNTAAN LIITTYVÄT TEKIJÄT
Jo seitsemänkymmentäluvulla havaittiin, että antamalla aiemmin NTD-lapsia synnyttäneille äideille runsaasti foolihappoa myöhemmän raskauden alkuvaiheessa saatiin sairaiden lapsien frekvenssi pienenemään jopa 70 prosentilla verrokkiryhmään verrattuna (kaksoissokkokoe), ja myös kokeet useilla NTD:ä ilmentävillä hiirimalleilla ovat osoittaneet, että paitsi foolihappo, myös metioniini ja myo-inositoli auttavat ehkäisemään häiriöiden muodostumista. Tämä on johtanut siihen että muunmuassa aamiaismuroihin lisätään USA:ssa foolihappoa, vaikka sen sivuvaikutuksena saattaakin olla tietyn vanhemmilla ihmisillä esiintyvän anemiamuodon peittäminen. NTD:ien ei ole kuitenkaa voitu osoittaa johtuvan foolihapon puutteesta, toisin kuin aiemmin oletettiin, eikä tavallisilla päiväannoksilla ole havaittu merkittävää riskiä vähentävää vaikutusta. Mutta mikä on folaatin ja muiden NTD:itä ehkäisevien aineiden vaikutustapa? Ainakin on selvää että folaatti on olennainen osa monia tärkeitä aineenvaihdunnan reaktioita, kuten pyrimidiinien synteesiä, mikä johtaa siihen että folaatin puutteesta kärsivät useimmiten eniten juuri ne solut, joiden jakautuminen, ja sitä myöten DNA:n ja pyrimidiinien synteesi, on kiivasta. Koska NTD-raskauksien aikana ja jälkeen ei ole havaittu äitien veressä folaatin puutosoireita, todennäköisempi vaikutuskohde onkin kehittyvässä alkiossa olevat geneettiset folaatti-aineenvaihdunnan häiriöt. NTD-lasta kantavien äitien verestä ja lapsivedestä on havaittu normaalia suurempia homokysteiinipitoisuuksia, jonka on kanan alkioilla suoritetuissa kokeissa havaittu lisäävän NTD;n todennäköisyyttä ja vaikka vastaavuutta ei olekaan havaittu ainakaan eräässä rottakokeessa, ovat folaattiaineenvaihdunnan homokysteiinin hajottamiseen liittyvät tekijät tehokkaan tarkastelun kohteena. Yksi kandidaateista on metioniinisyntaasi, joka katalysoi 5-tetrahydrofolaatin ja homokysteiinin reaktion, joka tuottaa metioniinia ja tetrahydrofolaattia. Toinen entsyymi on 5,10metyleenitetrahydrofolaattireduktaasi (MTHFR). Ja kolmas on Cystationi beta-syntaasi. Ainoastaan MTHFR;stä on havaittu termolabiili alleelimuoto, joka näyttäisi laajan tilastollisen analyysin mukaan olevan hienoisesti yleisempi NTD:tä kantavilla yksilöillä. . Tällä hetkellä ei siis ole olemassa mitään erityisen vahvaa teoriaa folaatin vaikutustavasta. Folaatille resistenttejä muotoja on jäljelle jäävät kolmekymmentä prosenttia, joskin osaa näistäkin saattaisi pystyä torjumaan myo-inositolin (joissakin yhteyksissä b-vitamiineihin luettu aine) lisäämisellä ravintoon, mihin viittaisi se että curly tail- hiirillä, joille ominainen selkäydinkohju on resistentti folaatille ja metioniinille, vaivan ilmeneminen pienenee huomattavasti myo-inositolin lisäämisen avulla, ja että säädellyissä oloissa ilman tätä vitamiinia kasvavat normaalit rotan sikiöt kehittävät NTD:itä. Myo-inositolin toimintatavasta on olemassa teoria, jonka mukaan se lisäisi proteiinikinaasi C:n aktiivisuutta.
Takaisin sivun alkuun
MUUNTYYPPISTEN GEENIEN AIHEUTTAMAT HERKKYYDET, MUODOT JA HIIRIMALLIT
Päätyökaluna neurulaation tutkimuksessa ovat erilaiset hiirimallit, joita on runsain määrin. Tähänastisissa tutkimuksissa yli 80 geenin on havaittu olevan jollakin tavalla kytköksissä hermostoputken sulkeutumisen häiriöihin, ja vaikka suurimmasta osasta havaintojen taustalla olevista geeneistä toimintaa neurulaatiossa on kartoitettu vasta vähäisesti, voidaan osaa niistä pitää jo välttämättöminä normaalin neurulaation kululle. Näihin kuuluu ainakin planaarisen solu-polaarisuuden aineenvaihduntareitti, joka on oleellinen neuraaliputken sulkeutumisen indusoitumiselle, sekä SHH-geenin toiminta, joka säätelee neuraalilevyn taipumista neuraaliputkeksi. On myös varsin selvää, että monet muut solunjakautumiseen ja yksilönkehityksen kontrolliin liittyvistä geeneistä kuuluvat virheellisinä muotoina ollessaan näitäkin kehityshäiriöitä aiheuttavien joukkoon. Neurulaation kulkua sääteleviä geenejä ovat ainakin, erään artikkelin mukaan (2). transkriptiofaktori T, Kasvainsupressori BRCA1 ja PAX-perheen geenit.
6.-kromosomissa sijaitseva transkriptiofaktori T:n geeni koodaa 435 aminohapon proteiinia, jonka DNA:han sitoutuva osa on evolutionäärisesti erittäin stabiili ja sataprosenttisesti samanlainen hiirellä ja ihmisellä. On oletettu että proteiini on tärkeässä osassa ainakin mesodermin kehityksessä. Tästä geenistä löydettiin ihmiseltä kaksi polymorfista kohtaa, joista toisen, geenin intronissa numero 7. yksi kolmesta alleelimuodosta esiintyy useammin NTD:tä kantavilla yksilöillä, mikä tarkoittaa joko sitä, että alleelimuoto vaikuttaa vaihtoehtoisen m-RNA silmukoinnin välityksellä, tai on markkerina jollekin vielä tuntemattomalle polymorfismille.
BRCA1:n alleelimuodoista useita on yhdistetty moniin syöpiin ja geenin tarkoituksena on inhiboida epiteelisolujen erikoistumista ja otaksuttavasti myös signaloida neuroepiteelisolujen erikoistumista. Suoraa näyttöä geenin vaikutuksista ihmisiin ei ole mutta hiirimalleissa siinä olevien virheiden on havaittu johtavan NTD:ihin.
PAX-geeniperhe koodaa yksilönkehitykselle oleellisia transkriptiofaktoreita ja ne ovat evolutiivisesti konservoituneita. Näistä ennenkaikkea PAX1 ja PAX3 vaikuttaisivat olevan tekemisissä NTD:iden syntymisen kanssa. PAX3:n virheet tunnetaan useimmiten Waardenburgin syndrooman aiheuttajana, minkä laajoista oireista yksi on selkärankahalkiot. PAX1:n virheet aiheuttavat hermostoputken halkioita, mutta ilmeisesti vain lievintä muotoa spina bifida occultaa.
Takaisin sivun alkuun
VIITTEET
1.Nature Reviews Genetics 4, 784-793 2003. The genetic basis of mammalian neurulation. (2003)(Andrew J.Copp et al.)
2.Journal of Applied Genetics 43(3), 343-350 Genetic basis of neural tube defects. 1.Regulatory genes for the neurulation process. (Monika Gos, Agnieszka Szpecht-Potocka)
3.Current Opinion in Neurology. Prevention of neural tube defects: vitamins, enzymes and genes. (Copp, Andrew J.) C. Lippincot-Raven Publishers.
4.Neurobiology, Exploring the Brain. (Mark F.Bear, Barry W. Connors, Michael A. Paradiso) C. Lippincot Williams &Wilkins
5.Developmental Biology 210, 15-29 Notochord-Dependent Expression of MFH1 and PAX1 Cooperates to Maintain the Proliferation of Sclerotome Cells during the Vertebral Column Development.(Furumoto et al.)
Takaisin sivun alkuun